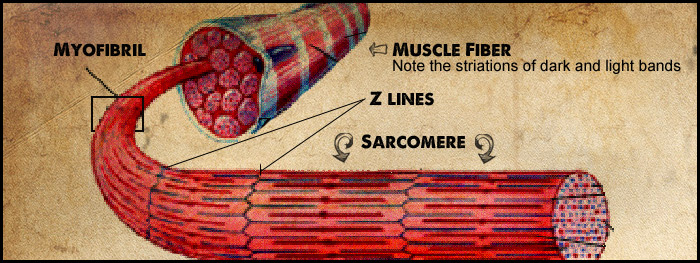
Review
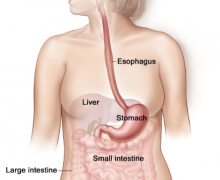
The digestive system plays an important role in the absorption of nutrients into the body. It takes the food we ingest, breaks it down mechanically and chemically in the mouth and stomach. It then absorbs nutrients, fats, proteins and water in the intestines before eliminating the waste through the rectum.
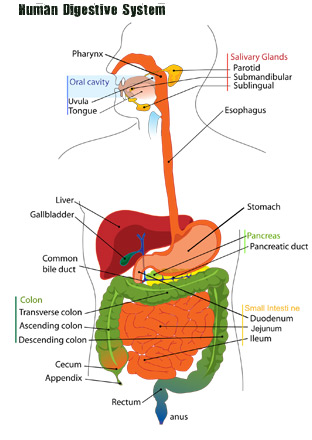
Major organs involved in the digestive system include the mouth, stomach, small intestine, large intestine, liver, pancreas.
The main steps in the digestive system
The digestive system is designed to do a few major things. They can be grouped into four categories:
- Ingestion
- Digestion
- Breakdown
- Excretion
Following Food from Mouth to Anus
To understand how our food is digested in the digestive system, it might be very useful to follow our food along its normal path, starting from the mouth.
Imagine for just a second that you’re hungry and your eyes gaze upon a nice home cooked thanksgiving turkey dinner. Your mouth starts to water. The salivary glands in your mouth are triggered to start producing saliva, a compound that will aid in the digestion of the meal.
As food enters your mouth, your teeth begin mechanically breaking down the food into small and smaller pieces. The saliva starts to chemically break it down as well. Soon, your conscious mind says, “lets swallow this food.” You swallow it and take another bite.
While you’re thinking about your next bite of food, your nervous system is helping to move the bolus (the food package you swallowed), down throat. A small flap of skin called your epiglottis makes sure your food goes down your esophagus. Movements of the smooth muscles, known as peristalsis help move that bolus down your esophagus. When it reaches your stomach, a sphincter opens and dumps the food in.
Inside the stomach, cells start to secrete different acids that help increase acidity to a pH of 2. This strong acidic environment kills most bacteria and starts to chemically break apart the food. Movements of the smooth muscles in the stomach, known as peristalsis mix and churn the food up more. After the food has been well mixed and has a consistency of oatmeal, it is ready to move to the small intestine. At this stage it is known as chyme.
To move into the small intestine, chyme must pass through the pyloric sphincter. From here it enters the duodenum, the first part of the small intestine. The liver mixes in bile, which helps break down fats in the food. The pancreas also secretes digestive enzymes that aid in digestion.
Most of the nutrients are absorbed from the small intestine and moved into the blood stream via a system of small folds, called vili.
After the food moves through the small intestine it enters the large intestine. The large intestine is named for the diameter of the cavity and not for the length. It is actually much shorter than the small intestine. The role of the large intestine is to remove any extra water from the digested material before it is finally excreted.
So there you have it – a basic rundown of what happens to the food we eat from the time we eat it, to when we excrete it. While we didn’t go into detail on all the steps, we would encourage you to explore as much as you can about each step of the process. Review our links below for more detailed explanations of the entire process.
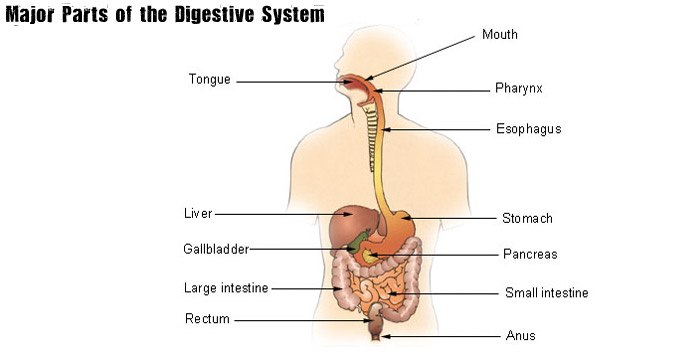
Digestive System Vocabulary Terms
-
- Anus: located at the end of the digestive system. The function of the anus is to expel feces, and unwanted semi-solid material produced during digestion.
- Appendix: a pouch like structure of the colon. The appendix is located near the junction of the small and large intestines. It is often referred to as the vermiform appendix or cecal appendix. The appendix is thought to be a vestigial structure in humans (an organ that has lost its original function). Today the appendix is prone to infection is often removed at the first sign of a problem.
- Ascending Colon: The large intestine can be divided up into different regions. The first section that takes digested material up is the ascending colon. It is smaller in caliber than the cecum, with which it is contiguous.
- Bile: yellowish, blue and green fluid secreted from the liver. Bile is sometimes called gall and is stored in the gallbladder between meals. It is secreted into the duodenum to aid int eh process of digestion of lipids via emulsification.
- Cecum: the beginning of the large intestine. The cecum is a pouch that connects the ileum with eh ascending colon.
- Chyme: the name given to the partially digested food that leaves the stomach via the pyloric valve into the small intestine (duodenum). Chyme, also known as chymus has the consistency of oatmeal.
- Descending Colon: the region of the large intestine that moves digested material downward through the left hypochondrium and lumbar regions.
- Duodenum: the first section of the small intestine. This hollow, jointed tube is only about 10-12 inches long. It connects the stomach to the jejunum and is where most of the chemical digestion takes place.
- Epiglottis: a small flap of elastic cartilage just below the pharynx that helps direct food down the esophogus and direct air into the lungs. Without the epiglottis, it would be difficult to
- Esophagus: a muscular tube that allows food to pass from the pharynx to the stomach. Through peristalsis the esophagus is able to push food from the pharynx to the stomach, even if the individual is standing on their head.
- Gall Bladder: a small organ that stores bile produced in the liver. It can hold about 50 mL of bile.
- Ileum: the last region of the small intestine. The ileum follows the other small intestine regions known as the duodenum and the jejunum. After the ileum, digestive material must pass through the ileocecal valve to get to the cecum.
- Jejunum: the middle section of the small intestine. It is preceded by the duodenum and followed by the ileum.
- Liver: a vital organ that produces chemicals necessary for digestion. In digestion it is extremely important in producing bile, which helps break down fats.
- Mouth: Often considered the first step in the digestive process. The chewing action of the mouth serves to mechanically break down food particles. Saliva produced here helps in chemical breakdown of food.
- Pancreas: an organ that aids in both the digestive system and the endocrine system. It aids in the digestive system by producing pancreatic juices, which contain digestive enzymes, which pass through the small intestine. These juices help break down carbohydrates, protien and fat in the chyme.
- Peristalsis: a radially symmetrical contraction of muscles that propagate down a tube. It occurs especially in the esophagus and the stomach. This movement helps the stomach mix up and break down our food.
- Rectum: the last portion of the large intestine. Human rectums are about 12cm long.
- Saliva: a substance produced by the salivary glands that aids in digestion. Human saliva is about 98% water. The other 2% consists of electrolytes, mucus, and antibacterial compounds.
- Salivary Glands: an exocrine gland that produces saliva, and amylase, among others.
- Sigmoid Colon: a loop of the large intestine about 40 cm long. It is the closest part of the large intestine to the rectum and anus.
- Stomach: a muscular organ in the gastrointestinal tract that helps churn up and digest food. The stomach lies between the esophagus and the small intestine. Smooth muscles in the stomach help to break down food mechanically via peristalsis.
- Transverse Colon: the longest and most movable part of the colon. It is the section of the large intestine just after the ascending colon and just before the descending colon.
Links to other Digestive System Resources
As you may have gathered from the above information, the digestive system is a complex and intricate part of the body. A number of things can go wrong with this system and when they do illnesses can be severe and lower a person’s quality of life significantly. You can learn more about these health issues and digestive system diseases at MyMed.com.












 Plant Topics
Plant Topics